Language comprehension is a fundamentally dynamic process, where the incoming speech stream is continuously segmented into a sequence of words and morphemes (Marslen-Wilson, 1973; 1975). This speech information interfaces with two joint but different neuro-cognitive processing systems: a left lateralised fronto-temporal system that is critical for grammatical language functions, and a distributed bi-hemispheric system that supports the broader semantic and pragmatic interpretation of the incoming utterance (Marslen-Wilson & Tyler, 2007; Tyler & Marslen-Wilson, 2008).
In our recent work we teased apart these different processing systems using functional neuroimaging (Bozic et al, 2010). Listeners heard either words that are specifically linguistically complex (words like prayed, which have the grammatical inflection ‘ed’), or words that make more general demands on the language processing system (words like claim, which have another word, clay, embedded in them). The results showed that activation for linguistically complex words is restricted to the left fronto-temporal regions. By contrast, words that are perceptually complex, due to increased competition between the whole word and the embedded word, activated a strongly bi-hemispheric set of regions, partially overlapping with the linguistic component.
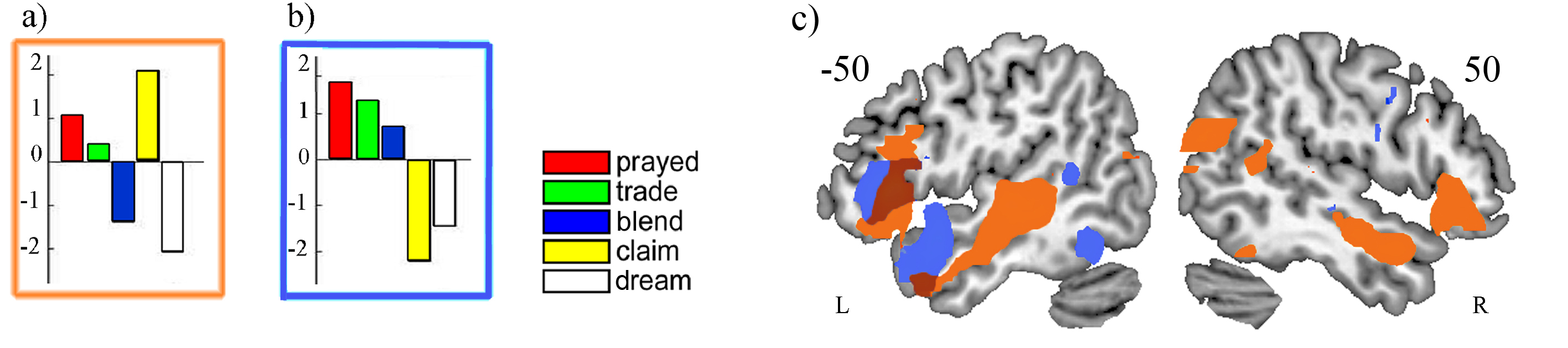
Figure 1: (a) MLM eigencomponent dissociating embedded from non-embedded words, (b) component dissociating linguistically compositional from non-compositional words, (c) overlay of the embeddedness (orange) and linguistic compositionality (blue).
Further results emphasise that only grammatical complexity (i.e. regular inflections in English) selectively activates the left lateralised fronto-temporal system. Other types of morpho-lexical complexity such as the lexicalised derivationally complex words (e.g. bravely, archer) do not trigger the selective engagement of the LH system. Instead, derived words robustly activate the distributed bi-hemispheric system, which supports the processing demands associated with mapping from sound to meaning. Multivariate RSA results show that this response is primarily driven by the properties of the derived form as a whole, with processing signatures reflecting the demands associated with separating the word from competing alternatives (Bozic et al, 2013; Bozic & Marslen-Wilson, 2010).
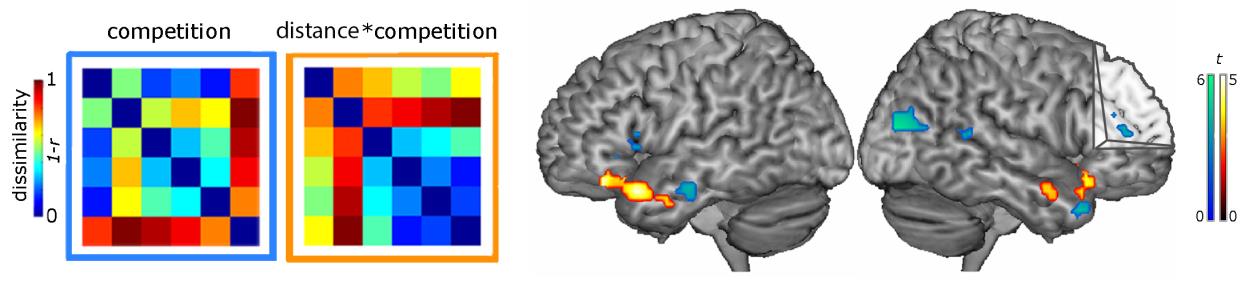
Figure 2: RSA models that capture perceptual competition and semantic relatedness x competition interaction, and areas of significant correlation for the competition effect (blue) and for the semantic relatedness x competition interaction (orange).
While there is strong evidence that LH system plays a critical role in supporting grammatical computations, data from brain damaged patients suggest that some aspects of grammatical processing could be supported bilaterally. A more specific characterisation of these mechanisms comes from our recent experiment that contrasted the processing of two types of grammatical sequences - inflectionally complex words (walks) and minimal phrases (I walk) - and contrasting them with grammatically simple words (walk). Results confirmed that the processing of inflected forms activates the LH system, emphasising its role in supporting complex grammatical computations. Critically however, they showed that the processing of simple linear phrases engaged bilateral temporal regions only, pointing to the capacity of bihemispheric system to support simple, linear grammatical computations (Bozic et al, 2011).
Prof Marslen-Wilson and Prof Tyler recently wrote a feature summarising research into the neuro-cognitive organisation of spoken language comprehension. You can read the full article here
Tracking the dynamics of language processing
The dynamic nature of language comprehension necessitates the use of techniques that can measure how and when the relevant processes emerge on a millisecond time scale. This is provided by magnetoencephalography (MEG) and electroencephalography (EEG). Our EMEG research (EEG and MEG combined) investigates the neural dynamics of both visual and spoken language comprehension, using several different approaches:
1) characterising the underlying networks at the source-level, computed with Minimum Norm Estimates
2) defining the inter-region communication, by looking at synchronisation in the signals between different areas (Lachaux et al., 1999)
3) investigating the neural computation in specific brain regions, by considering the source activity over a given region as a rich pattern of activity (Representational Similarity Analysis; Kriegeskorte et al., 2006).
Spoken words
Our fMRI research has shown a specific involvement of the left fronto-temporal system in processing combinatorial grammatical complexity (i.e., regular inflections, played). The source reconstruction of EMEG signals reveals a similar left-lateralised perisylvian network (Figure 2).
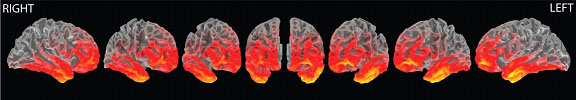
Figure 2: Mean source activity (N=17) for processing regular inflections, projected on an inflated brain. Yellow indicates more activity than red. Fronto-temporal activation is stronger on the left than on the right. This map correspond to an average of activity during the occurrence of the suffix [-ed].
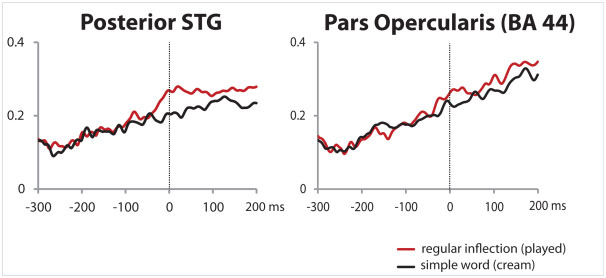
Figure 3: Time course of source activity for two selected regions of interest on the left hemisphere. Posterior STG (Superior Temporal Gyrus) is located on the temporal part of the brain and Pars Opercularis (BA44) is part of the frontal area. The comprehension of an inflected form (played, red curve) increased brain activity compared to a simple word (cream, black curve) during the time corresponding to the occurrence of the suffix (vertical dash line = 0 ms).
To define the inter-region communication and the specific timing of those interactions during the processing of inflected words, we analysed the oscillatory synchrony between specific brain regions. The results showed increased synchrony in the gamma band (20-60 Hz) between the left pSTG (posterior superior temporal gyrus) and the left pars opercularis (BA 44). This communication is highly specific to the regular past tense (played) and does not occur for simple words (cream). This cross-cortical communication occurs with specific timing, starting at 150ms before the acoustical burst of the suffix [-ed]. Our results suggest a specific cross-talk of fronto-temporal brain areas during the analysis of a linguistically complex word (Fonteneau et al., 2011).
References:
Marslen-Wilson, W.D. (1973). Linguistic structure and speech shadowing at very short latencies. Nature, 244, 522–523.
Marslen-Wilson, W.D. (1975). Sentence perception as an interactive parallel process. Science, 189, 226-228.
Maslen-Wilson, W.D. & Tyler, L. K. (2007). Morphology, language and the brain: the decompositional substrate for language comprehension. Philosophical Transactions of the Royal Society B, 362, 823-836.
Tyler, L.K. & Maslen-Wilson, W.D. (2008). Fronto-temporal brain systems supporting spoken language comprehension. Philosophical Transactions of the Royal Society B, 363, 1037-1054.
Bozic, M., Tyler, L.K., Ives, D.T., Randall, B., & Maslen-Wilson, W.D. (2010). Bihemispheric foundations for human speech comprehension. Proceedings of the National Academy of Sciences, 107, 17439-17444.
Bozic, M., & Marslen-Wilson, W.D. (2010). Neurocognitive contexts for morphological complexity: dissociating inflection and derivation. Language and Linguistic Compass, 11, 1063-1073.
Bozic, M., Tyler, L.K., Su, L., Wingfield, C., & Marslen-Wilson, W.D. (2013). Neurobiological systems for lexical representation and analysis in English. Journal of Cognitive Neuroscience, 25, 1678-1691
Bozic, M., Fonteneau, E., Tyler, L.K., Devereux, B., Buttery, P., & Marslen-Wilson W (2011). Grammatical categories in the fronto-temporal language network. Frontiers in Human Neuroscience, Conference Abstract: XI International Conference on Cognitive Neuroscience (ICON XI). doi: 10.3389/conf.fnhum.2011.207.00193
Fonteneau, E., Bozic, M. & Marslen-Wilson W. (2011). Cortical oscillations underlying morphological processing: an MEG/EEG study. Front. Hum. Neurosci. Conference Abstract: XI International Conference on Cognitive Neuroscience (ICON XI). doi: 10.3389/conf.fnhum.2011.207.00194.